What role does experience play in the development of vision?
The development of a structure as complex and sophisticated as the mammalian brain from simple embryonic beginnings is one of the most remarkable, and least understood, processes in biology. One particular aim of the many researchers working in this field is to determine the extent to which a person or animal's mind is 'hardwired' by genes, and the extent to which it is moulded by experiences – the inputs received by the maturing brain.
One function of the brain that has been extensively studied, in an attempt to answer this question, is the interpretation of visual information. This is an incredibly complex process, especially in primates (in the cortex alone, human vision is known to involve 32 separate areas, connected by no fewer than 187 neural pathways), and it is an area of neurology that has been the subject of extensive and ongoing research. In this essay, I will first provide a brief (and necessarily oversimplified) overview of the way in which the mammalian brain processes visual information, and I will then review the major discoveries that have been made about visual development. I will thus attempt to answer the question of whether "nature" or "nurture" is most significant in shaping, literally, the way we see the world.
The process of vision
The first stage of visual processing occurs in the retina of the eye, where layers of neurones refine and consolidate the input from the photoreceptor cells, before passing it down the optic nerve to the brain (Figure 1). At the optic chiasm, the neural pathways from each eye partially cross over, so that each hemisphere of the brain 'sees' only the contralateral (opposite) half of the field of vision.
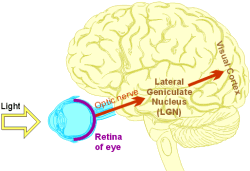
Figure 1: The main visual pathway in the human brain.
Several areas of the brain receive input from the eyes, including parts of the midbrain (those involved in eye movement control and 'fight or flight' responses) and the hypothalamus (where daylight patterns help to regulate Circadian rhythms). However, the most significant visual pathway – the one responsible for conscious perception – leads to the visual cortex, via an intermediate region in the thalamus called the lateral geniculate nucleus (LGN).
Afferent neurones from the LGN synapse in layer IV of the primary visual cortex (often referred to as area V1), where visual input is mapped topographically, with a particular area of cortex corresponding to a particular point on the retina. In layer IV, input from the two eyes is segregated into alternating strips. The cortical layers above and below layer IV consist of columns of cells that respond to specific characteristics of an image (Figure 2): ocular dominance columns compare the input from the two eyes, orientation columns respond to lines at specific angles, and cortical pegs are involved in colour vision. Pathways lead from V1 to specific areas of the cortex dedicated to the analysis of particular visual features, such as colour, form and motion. The final integration of all this information into a coherent interpretation of a scene appears to depend upon complex interactions between a large number of cortical areas working in parallel.
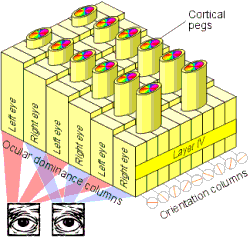
Figure 2: The organisation of area V1 of the visual cortex.
The role of experience
Experiments reveal that cells in area V1 of an 8 day-old kitten, whose eyes have not yet opened, show the same capacity for response to visual stimuli as adult cells do. It therefore appears that visual input is not required for the development of a functioning pathway from retina to visual cortex, a conclusion that is supported by studies in which mammals from various species have been reared in the dark, or had their eyelids sutured at birth, to produce binocular deprivation (a total absence of visual input from either eye). In such animals, the retina and LGN, and the pathways connecting them, were found to develop relatively normally. Abnormalities in the visual pathways leading from the retina can, however, be produced by genetic defects (such as albinism, which alters the way the optics nerves cross over at the optic chiasm, as illustrated in Figure 3).
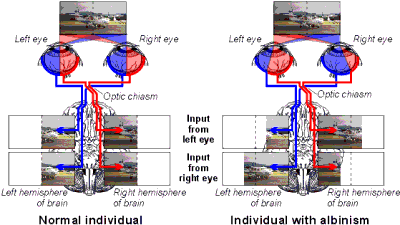
Figure 3: Comparison of the visual pathways in a normal individual (left) with those in an albino (right). Normally each hemisphere of the brain 'sees' only the contralateral half of the visual field, but in albinos, an abnormality in the development of the optic chiasm results in a part of the visual field in each eye being projected onto the ipsilateral hemisphere instead. (The dotted line on the photographs represents the midline.)
Within the visual cortex itself, binocular deprivation or the rearing of an animal in a limited visual environment can produce profound abnormalities. If kittens grow up wearing lenses that are designed so that only stripes of a certain orientation are brought into focus on the retina, a disproportionate percentage of the orientation columns in V1 are found to respond to lines of this particular orientation. Rearing of kittens in visual environments deprived of motion (by stroboscopic illumination), or deprived of 'form' (consisting simply of unpatterned dots) produces deficiencies in the areas of visual cortex that analyse these particular properties of a scene. Anatomical studies confirm that the development of connections within the visual cortex depends to some extent upon normal visual input. Injection of dye into the neurones of area V1 in a normal adult cat highlights characteristic 'clusters' of interlinked cells, but in animals that have been binocularly deprived during infancy, these clusters are not fully developed.
The interaction of the eyes
Counterintuitively, the effects on the cortex of depriving only one eye of vision (monocular deprivation) during development are even more profound than the effects of binocular deprivation. If one eye is sutured or blindfolded from birth, the strips of area V1 that receive input from that eye shrink, while those receiving input from the other eye enlarge correspondingly. This loss of cortical space by the deprived eye appears to be driven by competition from the nondeprived eye, rather than simply by atrophy of the unused areas, since it does not occur in the monocular region at the periphery of the visual field (where the two eyes do not compete). If vision in the deprived eye is restored in later life, recovery will be very limited – although the cortex recovers faster and more completely if the previously-normal eye is subsequently deprived. This suggests that the competition for cortical space from the 'good eye' continues to suppress input from the other eye even after vision in both eyes is restored.
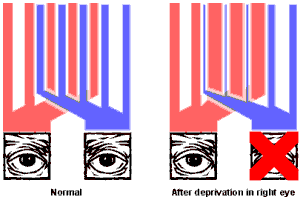
Figure 4: Illustration of the amount of cortical space in area V1 that is devoted to input from each eye, in normal individuals (left) and in those that have been monocularly deprived during development (right). The red bars represent the regions of V1 served by the left eye, and the blue bars represent the regions served by the right eye. Note that shrinkage of the regions receiving input from the deprived eye does not occur at the periphery of the visual field, where there is no competition from the non-deprived eye.
Normally, most of the cells in the visual cortex receive input to some extent from both left and right eyes. However, in individuals who develop amblyopia ('lazy eye') or strabismus ('cross eye') – conditions in which the images from the two eyes are incongruent – at a young age, an unusual number of cells are found that respond to only one eye or the other (as illustrated in Figure 5).
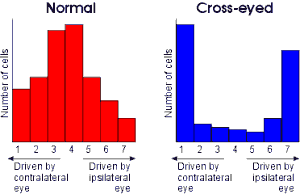
Figure 5: 'Ocular dominance histograms' for cells in the visual cortex of normal individuals (left) and those with strabismus (right). The numbers 1 to 7 are an arbitrary scale commonly used by neurologists to specify the extent to which a neurone is 'driven' by each eye: cells with a value of 1 respond entirely to input from the contralateral eye, those with a value of 7 respond entirely to input from the ipsilateral eye, and those with a value of 4 respond equally to input from either eye. Normally, the majority of cells respond to input from both eyes, but in individuals with strabismus (cross-eye), a condition that causes an abnormal mismatch in the images received by the two eyes, most cells are driven entirely by one eye or the other.
The critical period
In vision, as in many other aspects of neurology, there appears to be a critical period early in an animal's development during which its brain has a unique 'plasticity' and is particularly receptive to moulding by external stimuli. In every species so far studied, visual deprivation in adults appears to have little effect upon the cortex, but in infants of a certain age it can cause profound abnormalities. For example, as little as one hour of vision through striped lenses in 28 day-old kittens may be sufficient to bias the orientation columns of V1 to a particular orientation; this effect is not observed in adult cats. Monocular deprivation causes significant abnormalities in cats' brains only when it occurs at an age of between 3 and 16 weeks, and in humans, strabismus produces maximum effect upon cortical development at 2½ years of age (and no effect at all after the age of six). The exact neurological mechanism responsible for this period of plasticity during infancy is unknown, but it may involve the age-related changes that are known to occur in the density of certain neurotransmitter receptors on cells in the visual cortex.
Conclusion
In young, visually inexperienced animals, whose brains have never received visual input, it is possible to identify distinct areas of the visual cortex (albeit often in a crude and unrefined state), and an apparently functional pathway from retina to cortex. It can therefore be concluded that the fundamental layout of the visual processing areas in the brain is programmed by genes and not determined by external influences. However, experience clearly does play some role in constructing and refining the precise connections within the visual areas, particularly during infancy – a process that Jacobson (1978) dubs "functional validation".
It seems logical that genes and experience must both have a role to play in neural development. The complex functionality of the brain's visual areas could not possibly arise entirely out of undirected interactions with the environment in the absence of genetic influences, yet conversely, a mammal's genome could not possibly encode enough information to specify every individual connection within the brain. (Even if an organism's visual centres could be entirely 'hardwired', this would not be desirable, since some degree of flexibility is necessary in optimising the brain for the particular visual environment in which it finds itself, and in coping with ocular defects.)
Only by a combination of "nature" and "nurture" – with genes providing the basic blueprint and experience filling in the details – can a structure as complex as the mammalian brain develop to fulfil its maximum potential.
References
- Jacobson M (1978), Developmental Neurobiology
- Kolb B & I Q Whishaw (1996), Fundamentals of Human Neuropsychology
- Lam D M & C J Shatz (1991), Development of the Visual System
- Lund R D (1978), Development and Plasticity of the Brain
- Purves D (1988), Body and Brain: A Trophic Theory of Neural Connections
- Purves D & J W Lichtman (1985), Principles of Neural Development
- Shepherd G M (1994), Neurobiology
- Zeki S (1993), A Vision of the Brain
This was originally written as a university biology essay